Tanatocenózy měkkýšů v archeologických objektech v Hostivaři (Praha)
Knovízská kultura
Metodický přístup k hodnocení měkkýších nálezů
Výsledky malakozoologické analýzy
Přehled zjištěných druhů měkkýšů
Poznámky k determinaci měkkýšů
Praha, leden 2003
Jedním z hlavních zdrojů informací o někdejším prostředí a přírodním dění jsou pozůstatky pravěkých živočichů a rostlin, tj. fosílie nacházené společně s archeologickými objekty. V kvartérních vrstvách vzniklých na pevnině jsou to jak zbytky rostlinné (uhlíky, pyl, části rostlinných těl i otisky), tak živočišné (především kosti obratlovců a schránky měkkýšů). Každá z těchto skupin fosílií podává svědectví o přírodě minulých dob. Tyto poznatky se zčásti překrývají, zčásti jsou pro každou z uvedených skupin charakteristické, takže se vzájemně doplňují. Z toho tedy vychází i jedno z důležitých doporučení, že pro získání co nejúplnějšího obrazu přírodního prostředí je nutno interdisciplinárního přístupu ve výzkumu.
Zatímco kostem, zejména velkých savců, se odedávna věnovala patřičná pozornost, byly méně nápadné měkkýší pozůstatky spíše opomíjeny a jejich pozornost se soustřeďovala jen na nálezy některých nápadných velkých druhů. Na menší druhy nebo zlomky schránek se obvykle nebral zřetel, ačkoliv právě jejich výpověď může být velmi cenná. První souhrnné dílo věnované účelu hodnocení přírodního prostředí na základě měkkýších pozůstatků pochází z Velké Británie (Evans 1972), kde byla role měkkýšů jako význačných paleoenvironmentálních indikátorů dosti podrobně zpracována. V rámci středoevropských poměrů se touto problematikou zabývali také Ložek (1967) a Falkner (1969). V současnosti by již tedy mělo být samozřejmostí využívat fosilních měkkýšů při rekonstrukcích paleoenvironmentu, neboť znalost této skupiny bezobratlých živočichů je na značně vysoké úrovni.
Limitující podmínkou zachování schránek a lastur měkkýšů je primární vápnitost zkoumaných sedimentů. Vápník, vázaný ve formě uhličitanu vápenatého (CaCO3), tak příznivě ovlivňuje půdní reakci, která je v takových případech zásaditá. V neutrálních až kyselých půdách, resp. sedimentech se schránky a lastury měkkýšů rychle rozpouštějí.
Seskupení ulit a lastur měkkýšů tvoří tzv. tanatocenózy, tj. společenstva neživých jedinců. Tanatocenózy sestávají z pozůstatků jak velkých jedinců, tak malých. Nutno však podotknout, že v převážné většině jsou ulity velkých jedinců rozbity na menší kousky, neboť odolávají tlaku půdního profilu méně, než ulity měkkýšů drobných rozměrů. Přesto lze i zlomky ulit velkých jedinců snadno determinovat, neboť každý druh je typický svými znaky, které nacházíme na jeho ulitách (konchologické znaky). Jedná se především o povrchové skulptury jako rýhování, žebírka a jejich četnost na určité ploše zlomku. Naopak i absence povrchových struktur u druhů, jejichž ulity jsou hladké (za živa většinou lesklé), není determinační překážkou, neboť lze tento znak kombinovat se zakřivením zlomku a jeho tloušťkou. Významným znakem jsou též jizvy po chlupech, např. u zástupců srstnatých plžů. Tito plži (namátkou zmiňujeme Trichia hispida, Trichia sericea, Urticicola umbrosus) se vyznačují tím, že po jejich uhynutí zůstává v místě chlupu drobná jizva v podobě dolíku, patrného až po zvětšení pod stereomikroskopem. Vzájemné postavení jednotlivých jizev vůči sobě, jako jsou vzdálenost a prostorové uspořádání, pak umožňuje determinaci. Velmi důležitými určovacími znaky jsou také intrakonchologické struktury – zuby, lišty a lamely, které se nacházejí v ústí a v hlubších partiích závitů ulit (např. někteří zástupci čeledí Vertiginidae, Chondrinidae, Orculidae, Pupillidae, Buliminidae, Clausiliidae, Hygromiidae a dalších). Máme-li pak zlomek ulity s takovouto dochovanou strukturou, lze určit, o jaký druh se jedná. Dlužno upozornit, že ve všech těchto případech hraje při determinaci významnou úlohu zručnost a cvik determinátora.
Vzhledem k nízké pohyblivosti měkkýšů a jejich úzké vazbě na substrát můžeme při studiu jejich stanovištních nároků vycházet z podobných principů jako při studiu rostlinných společenstev. Živí měkkýši vytvářejí většinou ekologicky vyhraněná společenstva (malakocenózy), která jsou vázána na určitá stanoviště. Rozeznáváme tak například lesní, luční, stepní, mokřadní, vodní malakocenózy. V rámci těchto velkých skupin lze podrobněji dělit malakocenózy do dalších podskupin. Ve stručném podání to znamená rozdělení těchto druhů do ekoskupin, kam zařazujeme měkkýše stejných ekologických charakteristik – kombinujeme přitom jejich filii například k uzavřeným plášťovým formacím (lesní stanoviště), k otevřeným biotopům (stepní, luční stanoviště), ve vodním prostředí pak vazbu na tekoucí a stojaté vody, na výšku vodního sloupce i přítomnost vodních makrofyt. U suchozemských měkkýšů zároveň posuzujeme vazbu na mikroklimatické poměry jejich stanovišť, takže například v rámci lesních druhů můžeme dále odlišit ekoskupinu druhů lužních lesů (Macrogastra ventricosa, Clausilia pumila), ekoskupinu druhů teplejších prosvětlených lesů a hájů (Fruticicola fruticum, Helix pomatia) atd.
Pro suchozemské měkkýše je důležitá povaha půdy, resp. druh půdního typu. Určité druhy, např. Xerolenta obvia, Chondrula tridens, do jisté míry i Pupilla muscorum a Cecilioides acicula, zřetelně dávají přednost podkladu sedimentů nezpevněných před zpevněnými. Tyto prvky nazýváme jako pedofilní, protikladem jsou jim prvky petrofilní, vázané na zpevněné sedimenty.
Kombinací všech ekologických nároků jednotlivých měkkýších druhů byly vyčleněny 4 hlavní ekologické skupiny s 10 podskupinami dále ještě vnitřně členěnými (Ložek 1964).
Pevnost ekologické vazby měkkýších druhů na určitá stanoviště mohou kolísat v závislosti na nadmořské výšce a geologické a geomorfologické pestrosti daného území. V daných podmínkách se proto některé druhy mohou mírně odlišně chovat mimo souvislý areál svého výskytu. Velkou roli v tom mohou hrát také mikroklimatické podmínky, takže i v nižších polohách můžeme najít druhy typické pro montánní a submontánní stupeň – zde se jedná o výskyty v inverzních polohách (hluboce zařízlá údolí vodních toků, podmrzlé sutě)
V případě hodnocení měkkýších pozůstatků v archeologických objektech často dochází k jevu, který je zcela výjimečný, ne-li dokonce vyloučený ve volné přírodě. Jedná se o výskyty takových druhů, které ve zjištěné tanatocenóze zcela vybočují a již na první pohled nezapadají do obvyklé malakocenózy. Příkladem mohou být četné nálezy vodních druhů (např. velcí mlži) v tanatocenóze společně se suchozemskými (lesními, stepními apod.). Zde je nutno k analýze přistupovat velice opatrně, neboť je zřejmé, že ostatky těchto mlžů tvoří cizorodou složku, která byla druhotně přimíšena do původní terestrické malakocenózy. Tento jev se často týká odpadních jam, kde koncentrace schránek velkých mlžů může dosahovat až několika stovek nebo tisíců. Naopak, máme-li v tanatocenóze převažující vodní druhy, obvykle s určitým podílem drobných vodních zástupců, a méně zastoupené druhy suchozemské, můžeme uvažovat sedimentaci ve vodním prostředí (např. vodní příkopy), kam byly suchozemské druhy druhotně spláchnuty z příbřežních poloh. Velice těžko si lze představit, že by si tehdejší člověk povšiml vodních měkkýšů milimetrových rozměrů, které mohl přemístit na souš a kontaminovat původní suchozemské malakocenózy.
Tanatocenózy měkkýšů vzhledem k výše vybraným případům rozdělujeme proto do dvou složek:
1. Složka autochtonní – taková složka, která je tvořena druhy vyskytujícími se na stanovišti primárně (in situ). Jedná se hlavně o nejmenší a středně velké druhy, u nichž můžeme vyloučit druhotné přemístění a přimíšení k původnímu společenstvu.
2. Složka allochtonní – složka cizorodá, která byla do původní autochtonní složky přimíšena druhotně (ve většině případů záměrně). V tomto případě můžeme za allochtonní složku označit nálezy například velkých mlžů v čistě terestrickém prostředí, do něhož byly přeneseny z některé blízké vodoteče nebo vodní nádrže. Za allochtonní složku můžeme ve zvláštních případech považovat také nálezy těch největších plžů – Helix pomatia (hlemýžď zahradní) a Cepaea spp. (páskovky). Obdobně jako u mlžů, kteří mohli sloužit jako potrava a zpestření jídelníčku, lze i u hlemýždě předpokládat občasnou konzumaci; upravený hlemýžď je považován za postní jídlo. Páskovky, které jsou vzhledem k velikosti hlemýždě mnohem menší, naopak mohly upoutávat svou pestře páskovanou ulitou a není vyloučeno, že se jejich ulity používaly jako hračky, ozdoby nebo dokonce jako nějaká forma platidla. Nikterak to však nesnižuje jejich výpovědní schopnosti a naopak mohou popisovat stav a vzhled i v širším okolí sídlišť, odkud mohly být transportovány.
Vedle těchto záměrných zavlečení člověkem může docházet k druhotné kontaminaci původních malakocenóz i činností zvířat (tzv. bioturbace). Drobní hlodavci i větší půdní bezobratlí mohou vrtáním chodeb promísit vrstevní sledy. Časté jsou případy, kdy i přímo měkkýši mohou druhotně kontaminovat původní společenstvo – například terikolní drobný slepý plž Cecilioides acicula aktivně proniká až do hloubky 180 cm (Evans 1972). Jedná se o pedobiontní druhy a takové je nutno z analýzy vyloučit.
Poměrně podrobně bylo na metodiku odběrů vzorků pro malakologické analýzy i hodnocení případů autochtonních a allochtonních složek poukázáno v některých pracích s mezioborovou tématikou (Ložek 1981, 1998).
K malakozoologické analýze bylo předloženo 1322 vzorků, v nichž bylo celkem determinováno 5335 jedinců patřící 23 měkkýším taxonům[1]. Mezi determinovanými druhy převažovaly suchozemští plži, jako jediný vodní zástupce byl zjištěn mlž Unio crassus (velevrub tupý). Přehled všech zjištěných druhů je uveden v následujícím seznamu.
Přehled zjištěných druhů měkkýšů
Nomenklatura a systém jsou uvedeny podle Juřičkové, Horsáka a Berana (2001). Za názvem druhu následuje zoogeografické rozšíření a ekologická charakteristika (Kerney, Cameron a Jungbluth 1983, Ložek 1964).
Třída: Gastropoda (plži)
Podtřída: Pulmonata (plicnatí plži)
Čeleď: Cochlicopidae – oblovkovití
1. Cochlicopa lubricella (Rossmässler, 1835) – oblovka drobná
Zoogeografické rozšíření: Holarktické
Ekologická charakteristika: Obývá slunné xerotermní biotopy, hlavně stepi různého druhu a xerotermní skály; nejhojnější a nejtypičtější je na vápencových skalách a krasových stepích, kde místy žije v ohromném množství. Proniká místy i na biotopy mezofilní, např. na středně vlhké louky, zastíněné skalky apod. většinou ve statnějších formách. Místy žije též na lesních sutích v opadu mezi balvany.
Ekologická skupina: 6.
2. Cochlicopa lubrica (O. F. Müller, 1774) – oblovka lesklá
Zoogeografické rozšíření: Holarktické
Ekologická charakteristika: Obývá biotopy různého druhu , hlavně středně vlhké až mokré. Nejhojnější je na údolních nivách, kde žije na lukách, v olšinách i na březích vod, též ve vlhčích svahových lesích, v zahradách apod.
Ekologická skupina: 7.
Čeleď: Pupillidae – zrnovkovití
3. Pupilla muscorum (Linnaeus, 1758) – zrnovka mechová: holarktický
Zoogeografické rozšíření: Holarktické
Ekologická charakteristika: Obývá travnaté, suché a slunné stráně, řidčeji teplé skály. Miluje zvláště hlinité stepní stráně na vápnitém podkladu (spraš, slíny), druhotně se objevuje na zříceninách, náspech, mezích apod. Vyhýbá se vlhkým zalesněným oblastem. Ekologická skupina: 5.
Čeleď: Valloniidae – údolníčkovití
4. Vallonia costata (O. F. Müller, 1774) – údolníček žebernatý
Zoogeografické rozšíření: Holarktické
Ekologická charakteristika: Drobný plž, který obývá především bylinné formace – suché teplé stráně, meze a xerotermní skály. Z druhotných stanovišť nejčastěji proniká na hradní zříceniny nebo do zahrad. Nezřídka bývá nalézán i na středně vlhkých biotopech, především v ekotonech (např. na okrajích zalesněných údolí). Je indikátorem jak suchých, tak středně vlhkých stanovišť.
Ekologická skupina: 5.
5. Vallonia pulchella (O. F. Müller 1774) – údolníček drobný
Zoogeografické rozšíření: Holarktické
Ekologická charakteristika: Žije jako předešlý druh, ale je méně vázán na vlhčí stanoviště. Lesním oblastem se přísně vyhýbá. Nejčastěji se vyskytuje na mezích, pastvinách, stepních stráních, teplých skalách a podobných stanovištích.
Ekologická skupina: 5.
6. Vallonia excentrica Sterki 1893 – údolníček šikmý
Zoogeografické rozšíření: Holarktické
Ekologická charakteristika: Plž blízce příbuzný druhu Vallonia pulchella, jehož systematické postavení však není dosud řádně vyjasněno. Vzhledem k malým znalostem o ekologických preferencích jsou údaje o jeho životních nárocích sporé. Nejčastěji se vyskytuje ve společnosti druhu Vallonia pulchella na suchých stanovištích. Může pronikat i na středně vlhká stanoviště, zde však není doprovázen shora uvedeným druhem. Na základě současných znalostí lze tento druh řadit mezi obyvatele otevřených suchých až středně vlhkých biotopů. Lesním oblastem se vyhýbá.
Ekologická skupina: 5.
Čeleď: Vertiginidae – vrkočovití
7. Truncatellina cylindrica (Férussac, 1807) – drobnička válcovitá
Zoogeografické rozšíření: Evropské
Ekologická charakteristika: Obývá slunné travnaté stráně a skály, jedná se o význačný xerotermní druh plže. Nikdy neproniká na vlhká stanoviště. Lesním biotopům se striktně vyhýbá.
Ekologická skupina: 5.
8. Vertigo pusilla O. F. Müller, 1774 – vrkoč lesní
Zoogeografické rozšíření: Evropské
Ekologická charakteristika: Význačný prvek lesní. Žije ve vlhkém listí a mechu, též pod zetlelými větvemi a kůrou nebo na stinných skalách. Přednost dává smíšeným porostům javorů a lip s jasanem a jilmem.
Ekologická skupina: 1.
9. Vertigo angustior Jeffreys, 1830 – vrkoč útlý
Zoogeografické rozšíření: Evropské
Ekologická charakteristika: Význačný zástupce vlhkých až podmáčených biotopů, snese i krátkodobé zaplavení. Dává přednost otevřeným biotopům, jako jsou vlhké louky, prameniště, otevřené plochy podél vodotečí a rybníků. Do zastíněných ploch neproniká a lesním biotopům se striktně vyhýbá. Plž je slabě kalcifilní.
Ekologická skupina: 9.
10. Vertigo pygmaea (Draparnaud, 1801) – vrkoč malinký
Zoogeografické rozšíření: Holarktické
Ekologická charakteristika: Plž typický pro otevřené biotopy, kde obývá bylinné formace od mokrých až po stepní a skalní stanoviště. Velmi hojně žije na prameništích, na sušších stanovištích je hojný v případě vyššího obsahu CaCO3 v půdě. Lesu se přísně vyhýbá.
Ekologická skupina: 5.
Čeleď: Buliminidae – hladovkovití
11. Chondrula tridens (O. F. Müller 1774) – trojzubka stepní
Zoogeografické rozšíření: Pontomeridionální
Ekologická charakteristika: Obývá slunné stepní stráně, řidčeji xerotermní skály v teplých oblastech. Význačný prvek stepní. Běžný druh našich xerotermních oblastí.
Ekologická skupina: 4.
Čeleď: Succineidae – jantarkovití
12. Succinella oblonga (Draparnaud, 1801) – jantarka podlouhlá
Zoogeografické rozšíření: Eurosibiřské
Ekologická charakteristika: Je silně vázána na vlhká prostředí. Obývá nejen břehy, nýbrž i vlhké louky, luhy, údolní olšiny, často i daleko od vody. Častěji proniká do plášťových formací, ale můžeme ji zastihnout i na otevřených biotopech.
Ekologická skupina: 8.
Čeleď: Ferussaciidae – bezočkovití
13. Cecilioides acicula (O. F. Müller 1774) – bezočka šídlovitá
Zoogeografické rozšíření: Meridionální
Ekologická charakteristika: Drobný plž, který je přizpůsobený podzemnímu životu. Tělo i šídlovitá ulita postrádají pigmentu a jsou sklovitě bezbarvé. Druhu chybějí oči. Žije terikolně v neodvápněných půdách stepních biotopů, druh často zastihneme i na skalních stepích. Aktivně proniká až do hloubky 180 cm (Evans 1972). Vyžaduje lehké, nepříliš jílovité půdy, vždy primárně vápnité.
Ekologická skupina: 4.
Čeleď: Vitrinidae – skleněnkovití
14. Vitrina pellucida (O. F. Müller, 1774) – skleněnka průsvitná
Zoogeografické rozšíření:Holarktické
Ekologická charakteristika: Obývá lesy, údolní porosty, břehy vodotečí, též xerotermní skály a stepní stráně (při dostatečném krytu), je běžná i v kulturní krajině. Z hlediska ekologických nároků se jedná o nenáročného plže široké ekologické valence.
Ekologická skupina: 7.
čeleď: Zonitidae – zemounovití
15. Vitrea contracta (Westerlund, 1871) – skelnička stažená
Zoogeografické rozšíření: Západopalearktické
Ekologická charakteristika: Obývá obvykle sušší stanoviště – lesnaté skalnaté stráně a lesní sutě. Terikolně žije i na stepních stráních a na xerotermních skalách, do půdního profilu však neproniká tak hluboko jako Cecilioides acicula. Vyžaduje provzdušněné, lehké půdy s vyšším obsahem CaCO3.
Ekologická skupina: 7.
čeleď: Bradybaenidae – keřovkovití
16. Fruticicola fruticum (O. F. Müller, 1774) – keřovka plavá
Zoogeografické rozšíření: Evropské
Ekologická charakteristika: Obývá většinou háje a křoviště, v teplejších oblastech proniká také do luhů a lesů. Druhotně může pronikat do křovin na mezích a příkopů u silnic.
Ekologická skupina: 2.
čeleď: Hygromiidae – vlahovkovití
17. Euomphalia strigella (Draparnaud, 1801) – keřnatka vrásčitá
Zoogeografické rozšíření: Středoevropské
Ekologická charakteristika: Význačný prvek lesostepi; obývá xerotermní křoviště, stepní stráňky, skalní stepi, xerotermní háje. V chladnějších oblastech se jeví jako čistě stepní druh, v teplejších proniká běžně i do světlých suchých hájů.
Ekologická skupina: 6.
18. Trichia hispida (Linnaeus, 1758)
Zoogeografické rozšíření: Evropské
Ekologická charakteristika: Obývá většinou porosty vlhkých údolí, zvláště luhy, olšiny i vlhké louky, řidčeji se vyskytuje v lesích. Často proniká i na druhotná stanoviště, jako jsou zahrady, sady, při starých zdech. Stepním a xerotermním biotopům se většinou vyhýbá.
Ekologická skupina: 7.
19. Xerolenta obvia (Menke, 1828) – suchomilka obecná
Zoogeografické rozšíření: Jihovýchodoevropské
Ekologická charakteristika: Obývá suché stepní stráně, pole, meze; druhotně železniční náspy, příkopy, lomy. V chladnějších lesnatých oblastech se omezuje jen na vápencový podklad.
Ekologická skupina: 4.
Čeleď: Helicidae – hlemýžďovití
20. Cepaea vindobonensis (A. Férussac, 1821) – páskovka žíhaná
Zoogeografické rozšíření: Jihovýchodoevropské
Ekologická charakteristika: Obývá stepní stráně, xerotermní skály, křoviště i lesostepní formace. Často se objevuje na druhotných stanovištích – náspy, vinice, lomy, příkopy.
Ekologická skupina: 4.
21. Cepaea hortensis (O.F.Müller, 1774) – páskovka keřová
Zoogeografické rozšíření: Západo-středoevropské
Ekologická charakteristika: Obývá vlhčí místa v lesích, hájích a luzích, na úpatí lesních skal. Častá je v křovinách v úvozech a silničních zářezech. S oblibou proniká i do kulturních poloh – zahrady, sady, při starých zdech.
Ekologická skupina: 2.
22. Helix pomatia Linnaeus, 1758 – hlemýžď zahradní
Zoogeografické rozšíření: Středoevropsko-balkánské
Ekologická charakteristika: Obývá světlé háje a křoviště, hlavně v nižších teplých polohách. Častý je i plochách kulturních. Dává přednost vápnitému podkladu. Běžný prvek teplejších poloh.
Ekologická skupina: 2.
Třída: Bivalvia (mlži)
Čeleď: Unionidae – velevrubovití
23. Unio crassus Philipsson, 1788 – velevrub tupý
Zoogeografické rozšíření: Evropské
Ekologická charakteristika: Představitel sladkovodních zástupců velkých měkkýšů. Je striktně vázán na tekoucí vody, kde vyhledává stanoviště s písčito-bahnitým dnem. Je výrazně citlivý ke znečištění a v současné době je ohrožen vyhynutím. V minulosti býval hlavně v oblasti Polabí naším nejhojnějším velevrubem.
Ekologická skupina: 10.
Všechny determinované taxony jsou přehledně shrnuty v tabulce (Tabulka 1), z níž lze zároveň čerpat jak ekologickou charakteristiku všech druhů zařazených do ekologických skupin, tak i počet jedinců jednotlivých druhů.
Všechny terestrické druhy měkkýšů bylo možno determinovat přímo do druhu. Vedle těchto byly determinovány některé zlomky lastur pouze do rodu (rod Unio – velevrub), popř. do čeledi (č. Unionidae – velevrubovití). V těchto případech a vzhledem k potvrzenému výskytu mlže Unio crassus se lze domnívat, že zlomky s největší pravděpodobností patřily výše zmiňovanému druhu, nicméně pro korektnost je nutné uvažovat částečnou determinaci tak, jak ji umožňují zlomky lastur (viz. Tabulka 1).
Při determinaci pozůstatků měkkýšů byly zjištěny některé druhy, které je nutno vyloučit z následné paleoenvironmentální analýzy. Jedná se o ty jedince, kteří vykazovali známky recentního původu jako v případě plže Trichia hispida (3 jedinci), a dále o terikolní (pedobiontní) plže Cecilioides acicula a Vitrea contracta, kteří aktivně pronikají do půdního horizontu a mohou tak kontaminovat původní malakocenózy (C. acicula – 4302 jedinci, V. contracta – 2 jedinci). Po této korekci je vhodných celkem 1028 jedinců měkkýšů (Tabulka 1).
Tabulka 1: Kvalitativní a kvantitativní zastoupení determinovaných taxonů z archeologického naleziště v Hostivaři (Praha).
|
Hlavní ekologická skupina |
Vedlejší ekoskupina |
Druh |
Počet jedinců |
Počet jedinců vhodných k analýze |
||
|
A |
Les (všeobecně) |
1 |
W |
Vertigo pusilla |
1 |
1 |
|
2 |
W(M) |
Cepaea hortensis |
1 |
1 |
||
|
W(S) |
Fruticicola fruticum |
2 |
2 |
|||
|
Helix pomatia |
1 |
1 |
||||
|
B |
Bezlesí (všeobecně) |
4 |
S |
Cecilioides acicula |
(4302) |
0 |
|
Chondrula tridens |
6 |
6 |
||||
|
Xerolenta obvia |
2 |
2 |
||||
|
S(W) |
Cepaea vindobonensis |
2 |
2 |
|||
|
5 |
O |
Pupilla muscorum |
4 |
4 |
||
|
Truncatellina cylindrica |
130 |
130 |
||||
|
Vallonia costata |
584 |
584 |
||||
|
Vallonia excentrica |
25 |
25 |
||||
|
Vallonia pulchella |
207 |
207 |
||||
|
Vertigo pygmaea |
8 |
8 |
||||
|
C |
Les/bezlesí |
6 |
Ws |
Euomphalia strigella |
2 |
2 |
|
X |
Cochlicopa lubricella |
2 |
2 |
|||
|
7 |
M |
Cochlicopa lubrica |
4 |
4 |
||
|
Trichia hispida |
2+(3) |
2 |
||||
|
Vitrea contracta |
(2) |
0 |
||||
|
Vitrina pellucida |
1 |
1 |
||||
|
8 |
H |
Succinella oblonga |
1 |
1 |
||
|
D |
Mokřady Vodní biotopy |
9 |
P |
Vertigo angustior |
1 |
1 |
|
10 |
F |
Unio crassus |
23 |
23 |
||
|
Unio sp. frgm. |
17 |
17 |
||||
|
Unionidae frgm. |
2 |
2 |
||||
|
Počet celkem |
5335 |
1028 |
||||
Vysvětlivky:
Hlavní ekologické skupiny: A – les (všeobecně), B – bezlesí (všeobecně), C – les/bezlesí, D – mokřadní stanoviště, vodní biotopy.
Vedlejší ekologické skupiny: 1W – zapojený les; 2 – převážně les, 2W(M) – les až mezická stanoviště různého druhu, 2W(S) – les až suchá otevřená stanoviště; 4 – step až xerotermní stanoviště, 4S – step v užším smyslu, 4S(W) – zčásti zastíněná; 5O – otevřená stanoviště všeobecně, vlhké louky až stepi; lesy i bezlesí; 6 – převážně suché, 6Ws – zčásti zastíněné, 6X – převážně suché; 7- středně nebo různě vlhké, 7M – všeobecně; 8H – převážně vlhké; 9P – mokřady, břehy; 10F – tekoucí vody.
Pozn.: číslo v závorce značí počet jedinců druhotného původu, zkratka frgm. je fragment.
Nebudeme-li uvažovat ty druhy, které druhotně mohly kontaminovat vzorky s původními malakocenózami, je z Tabulky 1 dobře patrné, že lesní druhy byly ve vzorcích zastoupeny 4 druhy, druhů otevřených stanovišť bylo 9 a druhů indiferentních 6. Z mokřadních druhů byl ve vzorcích zachycen pouze 1 druh. Mezi vodními zástupci byl zjištěn 1 druh mlže.
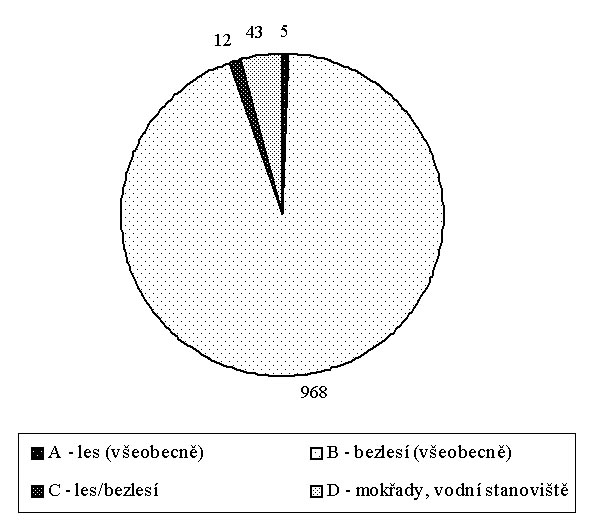
Z hlediska ekologických nároků byly zjištěny druhy všech hlavních ekologických skupin ve smyslu Ložka (1964) – lesní druhy, druhy otevřených stanovišť, indiferentní druhy i druhy mokřadní a vodní. Zatímco druhy otevřených stanovišť výrazně převažovaly, druhy lesní a druhy indiferentní spolu s mokřadními a vodními druhy byly zastoupeny spoře, jak si lze všimnout i na tabulkovém a grafickém zpracování poměrného zastoupení jedinců v hlavních ekologických skupinách (Obrázek 1).
Obrázek 1: Poměrné zastoupení jedinců v hlavních ekologických skupinách.
Hlavní ekologické skupiny |
Počet jedinců |
v procentech |
|
A – les (všeobecně) |
5 |
0,49 % |
|
B – bezlesí (všeobecně) |
968 |
94,16 % |
|
C – les/bezlesí |
12 |
1,17 % |
|
D – mokřady, vodní stanoviště |
43 |
4,18 % |
Mezi lesními druhy zaujímá výjimečné postavení drobný plž Vertigo pusilla. Tento plž žije v zapojených lesních celcích, kdy dává přednost vlhčím listnatým lesům s bujným bylinným patrem. Vzhledem k rozměrům jeho ulity (výška 2 mm, šířka 1,1 mm) lze s naprostou jistotou vyloučit jeho úmyslné zavlečení člověkem. Jak vyplývá z Tabulky 1 byl zjištěn pouze v jednom jedinci. Obdobně chudě zastoupeny byly další druhy lesních plžů – Cepaea hortensis, Fruticicola fruticum a největší náš zástupce Helix pomatia. Zatímco Cepaea hortensis dává přednost vlhčím lesním celkům a nezřídka proniká i na mezická stanoviště a do kulturní krajiny, plži Fruticicola fruticum a Helix pomatia dávají přednost prosvětleným lesním celkům suššího rázu. V nejteplejších polohách mohou pronikat také do zapojených lesů a luhů.
Zastoupení lesních druhů v analyzovaných vzorcích je velice chudé (0,49 %), nicméně spolehlivě indikuje v blízkosti sídliště přítomnost lesa charakteru mezických lesních stanovišť, které se střídaly s plochami vlhčími.
Ve vzorcích bylo celkem determinováno 968 jedinců náležející 9 druhům plžů, vesměs obdobných ekologických nároků. Celkově představují 94,16 %-ní podíl z počtu všech zjištěných jedinců. Převážnou většinu tvoří druhy malých rozměrů o několika milimetrech, u nichž lze předpokládat jejich původní výskyt na stanovištích (Truncatellina cylindrica, zástupci rodu Vallonia, Vertigo pygmaea). Jejich mobilita na větší vzdálenost je výrazně omezena a spíše ji lze odhadovat pouze v okruhu několika metrů.
Dále zjištění plži obdobných ekologických nároků, avšak mnohem větších rozměrů, byli xerotermní Cepaea vindobonensis a stepní Chondrula tridens spolu s Xerolenta obvia. Zatímco prvý jmenovaný dává přednost xerotermním křovinatým stanovištím, suchomilní Ch. tridens a X. obvia indikují stepní formace, přičemž u suchomilky X. obvia jde především o vysokostébelná travní společenstva. Je nutno podotknout, že se nemusí jednat o step v pravém slova smyslu, podmíněnou klimaticky a edaficky, ale i různé antropicky podmíněné krátko- a vysokostébelné travní formace.
Mezi tyto druhy řadíme měkkýše hlavní ekologické skupiny C s vedlejšími ekoskupinami 6, 7 a 8. Celkem jich bylo zjištěno 6 v souhrnném počtu 7 jedinců, což představuje 1,17 %-ní podíl v celkovém kvantitativním zastoupení.
Druhy indiferentní nelze v paleoenvironmentální analýze příliš využít, nicméně tvoří doprovodnou složku výrazných indikačních druhů, jak lesních, tak druhů otevřených stanovišť. V případě středně velkého plže Euomphalia strigella se jedná o doklad xerotermních křovinatých stanovišť, kterým dává výraznou přednost, zatímco na stanoviště vlhká nikdy nezasahuje. Naopak vlhkomilná Succinella oblonga svým výskytem dokládá přítomnost ploch vlhčího charakteru. Na výskyt indiferentních druhů měkkýšů je tedy nutno se dívat případ od případu.
Mokřadní druhy a druhy vodních stanovišť
Co se týče kvalitativního zastoupení, byly zaznamenány pouze dva druhy – mokřadní Vertigo angustior v počtu 1 jedince a vodní Unio crassus (23 jedinců). K nim je nutno zmínit další determinované zlomky lastur do rodu (Unio) a do čeledi (Unionidae), které s největší pravděpodobností náležejí druhu U. crassus. Celkově tyto druhy představují 4,18 % podílu v celkovém kvantitativním zastoupení.
Paleoenvironmentální rozbor[2]
Z ekologického rozboru zastoupení měkkýšů jednotlivých ekologických skupin vyplývá několik význačných rysů přírodního prostředí v areálu sídliště i v jeho těsném okolí.
Naprostá převaha druhů otevřených stanovišť, mezi nimiž se v rámci hlavní ekologické skupiny B vyskytuje i Cepaea vindobonensis vyžadující mírné zastínění, ukazují na souvislé otevřené plochy v rámci vlastního sídliště, antropicky ovlivněné, jejichž okraje nebo méně antropicky zatížené polohy (např. sešlapem, odstraňováním náletů keřů) přecházely v liniová společenstva s menšími plochami keřových porostů a enkláv stromů, jak můžeme usuzovat na základě občasného společného výskytu s keřovými, lesními a hájovými druhy. Toto lze doložit na základě velmi frekventovaného výskytu drobných plžů Truncatellina cylindrica, Vallonia costata, V. pulchella a V. excentrica, doprovázené drobným plžem Vertigo pygmaea, dávající přednost otevřeným polohám různého charakteru. Jako indikátor občasných keřových xerotermních formací zde vystupuje jednak ekologicky vyhraněná Cepaea vindobonensis a suchomilnější Euomphalia strigella. Vedle nich zaujímají důležité místo i druhy lesní a hájové jako drobný plž Vertigo pusilla, jež indikuje zapojený les, a velcí plži Helix pomatia a Fruticicola fruticum, vázaní spíše na stanoviště prosvětleného lesa a hájů, suššího charakteru. Tyto druhy řazené do hlavní ekologické skupiny A (lesní druhy všeobecně) byly sice zachyceny zcela ojediněle, nicméně s jejich výskytem v areálu sídliště je nutno počítat jako s důkazem, že i zde se již vyskytovaly plošky (patches) keřových a řídkých stromových porostů, jež za hranicemi sídliště pravděpodobně nabývaly větší rozlohy. Tento úsudek je zcela v souladu s pohledem na vzhled tehdejšího přírodního prostředí, neboť lze jen těžko usuzovat na to, že vlastní intravilán sídliště zachovával typicky lesní ráz. Ten je možno předpokládat v blízkém okolí sídliště, kde již přímý lidský impakt na vegetační a stromový kryt byl snížen, což umožnilo detekci lesních druhů sensu lato.
Ve vztahu k vlhkostním poměrům studovaného sídliště byla doložena přítomnost menších vlhčích nebo dokonce podmáčených ploch, jak to dokazují nálezy vlhkomilného plže Succinella oblonga a dokonce mokřadního Vertigo angustior. Tyto plochy byly jen fragmentárního charakteru a zcela potlačeny na úkor mohutného rozvoje ploch xerotermního rázu, jak to dokazuje naprostá převaha druhů ekologické skupiny B.
Indiferentní druhy mezických stanovišť (Cochlicopa lubrica, Trichia hispida a Vitrina pellucida) nebyly zastiženy příliš často a tvoří jen doprovodnou malakofaunu k převažujícím druhům otevřených stanovišť, méně pak druhům lesním.
Nálezy vodního měkkýše Unio crassus v areálu sídliště zasluhuje zcela samostatný přístup ve smyslu jeho vypovídací schopnosti. Jak bylo zjištěno, tento mlž nebyl doprovázen žádným jiným vodním druhem a naopak jeho společný výskyt s terestrickými druhy ukazuje na druhotné zavlečení lastur mlže do terestrického prostředí. V tomto případě je nutno jeho nálezy považovat jako allochtonní složku, která však nikterak nesnižuje jeho výpovědní hodnotu. Tento mlž je striktně vázán na tekoucí vody (rheofilní druh) a jeho výskyt v některém z nejbližších toků poblíž sídliště tedy ukazuje přímo na stav vodního toku. Ve srovnání s dneškem nejen z blízkého, ale i z rozlehlého území hostivařského sídliště znám není a je tedy zřejmé, že v době sídlištní aktivity byla kvalita vody nesrovnatelně vyšší než v současnosti.
Měkkýši ve vztahu k lidské činnosti
Jak bylo v úvodních kapitolách naznačeno, z hlediska výskytu měkkýšů v archeologických objektech je nutné posuzovat dvě základní hlediska – přirozený výskyt a druhotné zavlečení.
V případě přirozeného výskytu je zřejmé, že tehdejší lidé si jen těžko mohli povšimnout malých zástupců měkkýšů, jejichž velikost se pohybuje v rozmezí 1-4 mm. Lze u nich vyloučit úmyslné zavlečení, takže jejich výskyty vypovídají o vzhledu přírodního prostředí in situ. Jak bylo v ekologickém rozboru zmíněno, jedná se především o druhy otevřených stanovišť, k nimž se druží také drobný lesní druh Vertigo pusilla a vlhkomilnější Succinella oblonga a mokřadní Vertigo angustior. Na základě výskytů těchto měkkýšů lze rekonstruovat stanoviště in situ, jak již bylo výše zmíněno.
Druhotný výskyt některých měkkýších druhů je nutno posuzovat případ od případu. Ve vztahu k nálezům mlže Unio crassus je zřejmé, že se jedná o typickou allochtonní složku, tedy o měkkýše druhotně zavlečeného do archeologických objektů z některého blízkého toku. Zde se nabízí vysvětlení o využívání mlžů jako zdroje potravy. Nelze přesně doložit, zda tito mlži byli konzumováni pravidelně, naopak jejich slabý výskyt (23 jedinců U. crassus, 17 jedinců Unio sp., 2 jedinci Unionidae) ukazuje na občasnou konzumaci víceméně charakteru pouhého zpestřování si jídelníčku. Důležitou roli v tomto tvrzení hraje i početnost populace mlžů ve vodním toku, kde byli lidmi sbíráni. Nelze vyloučit, že opakovaným sběrem mlžů byla početnost populace dosti snížena, což se mohlo projevit i v nízkém početním zastoupení lastur v archeologických vrstvách. Případy, které byly zaznamenány na dolním toku řeky Labe, zcela jasně ukazují, že zde bylo využíváno velevrubů ke konzumaci masově, jak dokládají nálezy stovek až tisíců lastur v odpadních jamách. Takový případ se v sídlišti v Hostivaři nevyskytl, na druhou stranu jej nelze vyloučit.
Z hlediska druhotného zavlečení je možno uvažovat též výskyty velkých plžů Helix pomatia a Cepaea vindobonensis. Hlemýžď H. pomatia mohl být využíván k občasné konzumaci obdobně jako mlž Unio crassus. Byl však nalezen pouze v jediném exempláři, takže se jako více pravděpodobné nabízí jeho autochtonní původ v sídlišti.
Páskovky Cepaea vindobonensis a C. hortensis se vyznačují pestrobarevnými páskovanými ulitami, které jistě tehdejší lidi upoutaly svými výraznými barvami. Je tedy vysoce pravděpodobné, že mohly sloužit jako hračka, případně jako ozdoba nebo dokonce jako nějaká forma druhotného platidla, popř. výměnného artiklu. U ní vzhledem k mnohem menším rozměrům oproti hlemýždi nelze předpokládat sběr za účelem konzumace, neboť energetický výdaj s přípravou tohoto pokrmu by byl mnohem větší, než by člověk získal. Nicméně, ať se dostaly do objektu úmyslně nebo ne, což platí i v případe hlemýždě Helix pomatia, jejich vypovídací schopnost o paleoenvironmentu tím není nikterak snížena, neboť pokud byly úmyslně zavlečeny, vypovídají o širším okolí, v případě přirozeného výskytu v místě sedimentace vrstvy vypovídají přímo o místních podmínkách.
Cílem práce byla rekonstrukce přírodního prostředí pomocí struktury tanatocenóz měkkýšů. Z 1322 odebraných vzorků bylo determinováno 5335 jedinců patřící 23 druhům (22 terestrických, 1 vodní). Po vyloučení jedinců recentního původu a jedinců druhů terikolních bylo k analýze využito 1028 exemplářů. Ve vzorcích výrazně dominovaly druhy otevřených stanovišť, které indikují přítomnost bezlesí (pravděpodobně sekundární step, antropicky ovlivněná – Truncatellina cylindrica, Vallonia costata, V. pulchella, V. excentrica, Pupilla muscorum). Kromě nich byly zjištěny slabě zastoupené lesní a hájové druhy (Vertigo pusilla, Cepaea hortensis, Helix pomatia, Fruticicola fruticum) a druhy xerotermních keřových formací (Cepaea vindobonensis, Euomphalia strigella). Ojediněle byly zjištěny druhy vyžadující vyšší vlhkostní poměry stanovišť (Succinella oblonga, Vertigo angustior). Ze vzorků z terestrického prostředí byl získán větší počet lastur mlže Unio crassus, který pravděpodobně sloužil jako příležitostná potrava pro obyvatele sídliště.
Na základě zjištěných měkkýšů lze zrekonstruovat přírodní prostředí sídliště a jeho blízkého okolí vzhledu souvislých otevřených ploch v rámci vlastního intravilánu sídliště, jejichž okraje nebo méně antropicky zatížené polohy přecházely v liniová společenstva s menšími plochami keřových porostů a enkláv stromů, které za hranicemi sídliště nabývaly větší rozlohy.
Evans J. G. (1972): Land Snails in Archaeology. – Seminar Press, London and New York, 436 pp.
Falkner G. (1969): Die Bearbeitung ur- und frühgeschichtlicher Molluskenfunde. – Archäologie und Biologie, Forschungsberichte 15 (DFG), 112-140. Wiesbaden.
Juřičková L., Horsák M. & Beran L. (2001): Check-list of the molluscs (Mollusca) of the Czech Republic. – Acta Societatis Zoologicae Bohemiae, 65: 25–40. Praha.
Kerney M. P., Cameron R. A. D. & Jungbluth J. H. (1983): Die Landschnecken Nord- und Mitteleuropas. – Verlag Paul Parey: 1-384. Hamburg und Berlin.
Ložek V. (1964): Quartärmollusken der Tschechoslowakei. – Rozpravy Ústředního ústavu geologického, 31: 1-376. Praha.
Ložek V. (1967): Beiträge der Molluskenforschung zur prähistorischen Archäologie Mitteleuropas. – Zeitschrift für Archäologie, 1: 88-138. Berlin.
Ložek, V. (1981): Měkkýši v archeologii. – Archeologické rozhledy, XXXIII, 2: 166-175. Praha.
Ložek, V. (1998): Pozůstatky fauny v archeologických výkopech a jejich výpověď. Část I – základní údaje a měkkýši. – Archeologické rozhledy, L, 2: 436-451. Praha.
[1] Tabulkové zpracování všech determinovaných měkkýšů v jednotlivých vzorcích není vzhledem ke značné obsáhlosti údajů součástí této závěrečné zprávy a je odděleně přiloženo v elektronické podobě (formát .xls na disketě). V této zprávě je kladen důraz na ekologické charakteristiky jednotlivých taxonů a jejich vypovídací schopnost o vzhledu a stavu přírodního prostředí v oblasti sídliště a v jeho nejbližším okolí.
[2] V paleoenvironmentálním rozboru nebyly zohledněny druhy Cecilioides acicula a Vitrea contracta, kteří jako pedobiontní živočichové aktivně pronikají do půdního profilu a jejichž přítomnost v původních tanatocenózách, resp. malakocenózách je více než diskutabilní. Obdobně byly vyřazeny i nálezy 3 ulit plže Trichia hispida, které měly zřejmý recentní původ.